大学入試共通テスト「生物」第2回試行調査第5問A(遺伝子・11点配点)問題・解答・解説

解説
問1
少し長くなるが、この設問を解く上での基礎知識を順にまとめていく。すでに理解できている方は、理解できている部分は読み飛ばしてもよい。
【遺伝子組み換え技術】
遺伝子組み換え(genetic recombination)技術の基本を理解しておこう。遺伝子研究には、電気泳動・PCR(DNA増幅)など様々な技術があるが、問1で聞かれているプラスミド(plasmid)の役割を知っておこう。プラスミドの大腸菌への組み込みは、それ自体が本設問のような基礎研究の手法となるだけでなく、実際に医薬品生産として、大腸菌によるヒト・インスリン生産などで実用化されている。その実用の流れを知っておくと、その流れの一部を使った基礎実験の流れもわかりやすくなるはずである。
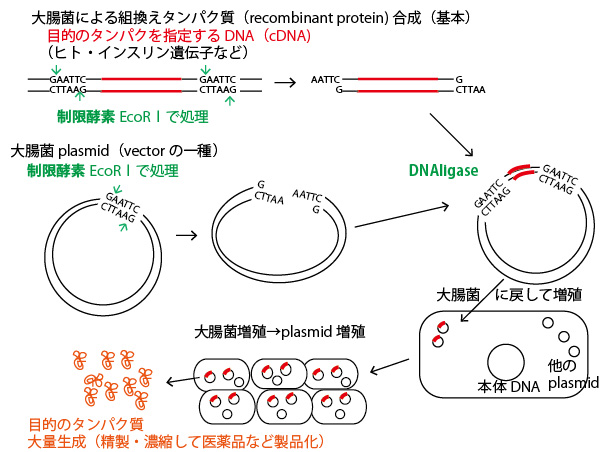
上記図のように、まず組み込みたい(医薬品生成などをしたい)遺伝子部分を含むDNA(実際はmRNAから逆転写酵素を使って作ったイントロン部分を含まないcDNA)を取り出す。赤がその遺伝子部分である。その遺伝子の両脇のDNAの特定の塩基配列を認識し切断する酵素(制限酵素restriction enzyme)を使って、DNA2本鎖を、一部、数塩基(ヌクレオチド)分(4塩基突出が多い)だけ一本鎖を突出させた形で切断する。(この図で使った制限酵素EcoRⅠではAATTが突出)
(なお制限酵素には様々な種類があり、認識配列や突出部分の塩基配列は種類により異なる。)
次に、大腸菌の中に本体DNAとは別個に複数存在する小型の環状DNA(プラスミド・plasmid)を取り出してきて、同じ制限酵素で切断する。するとプラスミドと目的遺伝子を含んだDNA断片が同じ突出DNA一本鎖を持つ。そこにDNAリガーゼ(ligase)というDNA2本鎖を連結する酵素を処理すると、AATTの突出部分同士が相補的に連結し、更に酵素の作用で切られていたヌクレオチドが献血され、目的遺伝子(赤い部分)を含むプラスミドができる。
それを大腸菌に戻して、その大腸菌を培養・増殖させると、大腸菌内でプラスミドも複製され、その中の遺伝子が発現し、目的のタンパク質が大量に生成される。それを生成して医薬品として使う。
【核酸(DNAとRNA)】
次に核酸(nucleic acid)についてまとめよう。核に存在する酸性の物質なのでそう命名された。酸性の性質を生み出しているのは鎖に規則的に存在するリン酸がH+を放出して電離している(マイナス電荷を帯びている)からである。
核酸にはDNAとRNAの2種があるが、その構造の違いを以下の図にまとめた。
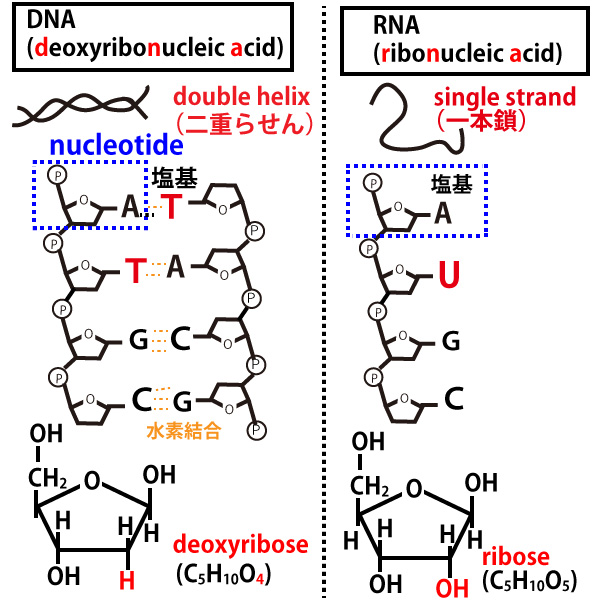
共通点は、青線で囲んだように「リン酸・糖・塩基」が結合してできているヌクレオチドを基礎単位とした鎖であることである。
一方、違いをまとめると以下のようになる
1、DNAは二本鎖(二重らせん)・RNAは一本鎖。
(注、ウイルスでは一本鎖DNA・二本鎖DNAという例外現象がある)
2、DNAの糖はデオキシリオボース(deoxyribose)、RNAの糖はリボース(ribose)で名前の由来となった。
(糖の基本は原子数はC:H:O=1:2:1である。たとえばグルコース(ブドウ糖)はC6H12O6である。炭素原子が5つの糖(五炭糖という)の基本形はリボースC5H10O5、その基本形から酸素原子を1つ取った(de、oxy)のがデオキシリボース(C5H10O4)と考えればよい)
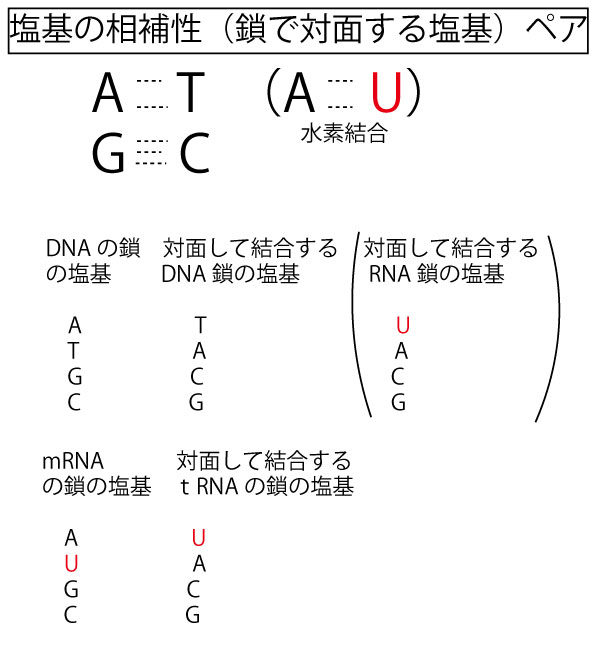
3、塩基の種類は、DNAはATGC、RNAはAUGC。
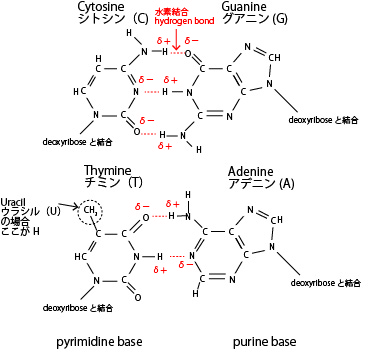
【塩基対(水素結合)のペア】
特定の塩基どうしは以下のように鍵と鍵穴のように水素結合(電気的な引き合いによる結合)をする。水素結合は共有結合より分離しやすく、これがDNA二重らせんをほどいて複製したり、RNAに転写する基礎となる。
そしてDNAが複製するときも、RNAに転写するときも、RNAどうしが結合するときも必ずもとの鎖の塩基と対面する塩基が決まっているこれを塩基の相補性という。
【DNA(RNA)の5´→3´方向と逆平行】
更にDNA・RNAの鎖の向きについて正確に知っておく必要がある。
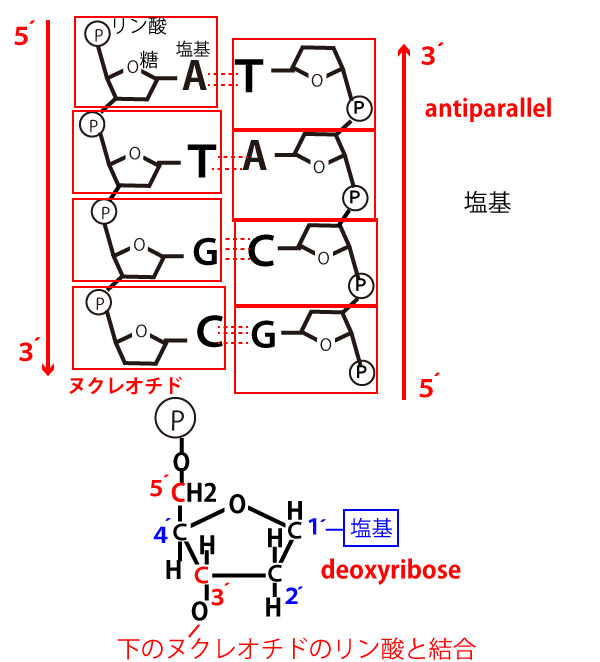
下図のように五角形の形をしたデオキシリボースの分子内の炭素原子には酸素原子(O)を上に置いて考えた場合、時計回りの順に「1´」「2´」「3´」「4´」「5´」の番号がついている。「1´」炭素は塩基と結合し、「3´」炭素は下側のヌクレオチドのリン酸と結合し、「5´」炭素は同じヌクレオチド内(上側)のリン酸と結合する。するとリン酸ー糖ーリン酸ー糖の繰り返しとなっている各鎖の外側には5´と3´の向きがあることがわかる。そして「5´→3´」の向きをDNAでもRNAでも鎖の正式な向きとする。これは日本語でも英語でも、横書きの一般の文章は「左→右」の向きに読むという約束があるのと同様、遺伝子(分子生物学)における文法だと考えてよい。
5´→3´方向に関して2つの重要な性質がある。
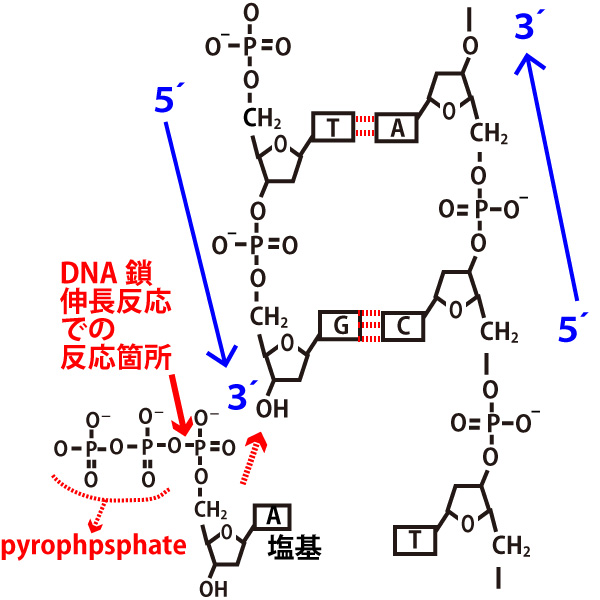
1、(DNAどうし、DNAとRNA、RNAどうしなど)鎖が対面する時、両方の鎖は5´→3´方向が逆向きになる。(antiparallel・逆平行)という。
2、DNAでもRNAでも鎖の伸長(合成)は5´→3´の方向に起きる。
右側の鎖を鋳型にして、左側のDNA鎖が伸長する場合の図で上記1.2を確認してほしい。
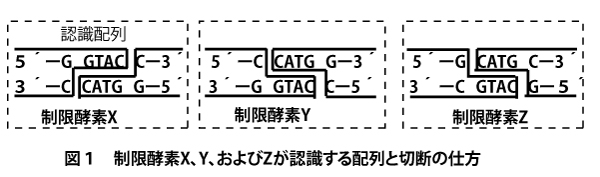
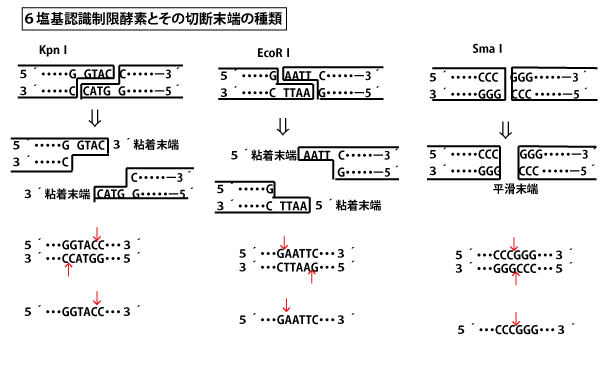
【制限酵素とその認識配列】
最初の「遺伝子組み換え技術」で出てきたように、DNAをある塩基配列(正確には塩基「対」配列)を認識して切断する酵素を制限酵素(reatriction enzyme)という。4塩基・5塩基・6塩基・8塩基など様々な長さを認識する酵素があるが、実験においても大学入試においても6塩基認識酵素が使われ出題されることが多い。
6塩基認識制限酵素の切断面は以下3通りがある。
まず、上記に代表例としてあげた3つの制限酵素が認識する配列の共通の特徴を見てほしい。
Kpn1の場合は、上側の鎖は5´ーGGTACCー3´で、下側の鎖を5´→3´方向に見ると5´ーGGTACCー3´で同じとなる。EcoR1の場合も5´ーGAATTCー3´は上下同じ、Sma1の場合も5´ーCCCGGGー3´は上下同じである。このように5´→3´方向に見ると両鎖の塩基配列が同じ並びになっている構想を「回文構造(palindrome)」という。日本語の「竹藪焼けた(タケヤブヤケタ)」、英語の「Madam ,I’m Adam.」のように右から読んでも左から読んでも同じになる文と同じである。
さて回文構造であることは同じであるが、切断方法が異なる。上下の鎖が対称的になるように切断するが、Kpn1などの場合は5塩基目のヌクレオチドと6塩基目のヌクレオチドの間(リン酸と糖の共有結合)を切断する。すると鎖が離れはじめ、2番目~5番目の塩基間の水素結合は切断される。そして、4塩基分の(ヌクレオチドの)1本鎖が突出して残される。この突出部は3´側であり、のちにこの突出部がDNAリガーゼにより他の同様の突出部との粘着の「のりしろ」となるので「3´粘着末端」という。
EcoR1などの場合は、1塩基目のヌクレオチドと2塩基目のヌクレオチドの間(リン酸と糖の共有結合)を切断する。すると鎖が離れはじめ、2番目~5番目の塩基間の水素結合は切断される。そして、4塩基分の(ヌクレオチドの)1本鎖が突出して残される。この突出部は5´側であり、のちにこの突出部がDNAリガーゼにより他の同様の突出部との粘着の「のりしろ」となるので「5´粘着末端」という。
一方Sma1にように上下とも3塩基目のヌクレオチドと4塩基目のヌクレオチドの間でスパッと切る「平滑末端」もある。しかし、これは、同タイプの制限酵素切断端とならどのタイプとでも結合してしまうので、特定の塩基配列の認識を重視したい実験の流れの中では使用されないことが多い(よって大学入試でもほとんど出題されない)
なお上図の下のように切断面に↓・↑、あるいは▽・△を書き込んで制限酵素の切断方法を示す。両鎖とも表記
することもあるが、更に簡略化すると「回文構造」であることを前提とすれば、上の鎖の6塩基に↓を書き込むだけで切断面を示すことができるので、学問的には、上の鎖の↓(あるいは▽)のみで示す。
【3´粘着末端と5´粘着末端は再結合しない】
3´粘着末端と5´粘着末端は4塩基の一本鎖が突出している点は共通である。しかし、両者が、接近・対面した時に5´→3´の方向(下図では→で示す)に並べると、一本鎖突出部分が同じ側で対面してしまい、塩基対を構成できないので結合ができない。(無理矢理、一本鎖突出部どうしが結合する形を考えた場合、連結部で鎖の向きが変わるので結合できない)。
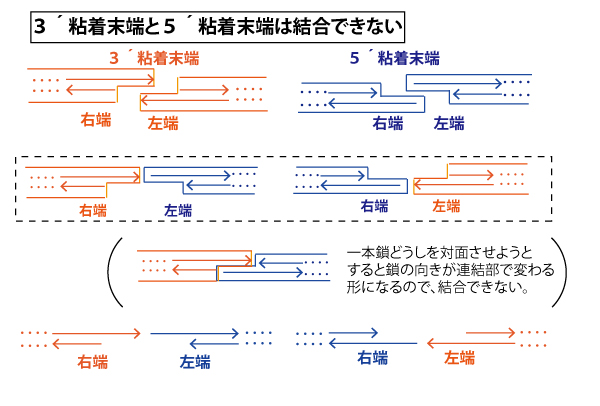
なお、図の上では、初学者にわかりやすいように鎖の枠を線で囲んだが、この囲み線を描くのに時間がかかるので、巣図の下のように→だけで表記する方向に慣れていこう。
【同じ制限酵素で切断した右端・左端は180°回転させると入れ替わる】
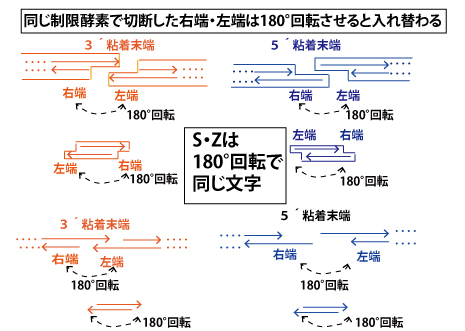
制限酵素の切断面に関し、平面上での図の説明でわかりやすいように、これまでは「左端」「右端」と説明してしたが、実際のマイクロチューブ(化学実験の試験管に相当する分子生物学実験でのチューブ)内では、空間的に浮遊しているので、右・左があるわけではないし、溶液内で浮遊する中で回転しながら位置も変わっていく。ただ空間的な回転まで表現することは困難なので、180°回転したことを考えてみよう。すると実は「回文構造」の性質から、「右端」と「左端」は相互に入れかわることができるとわかる。つまり突出部は平面上で「右」に描こうが「左」に描こうが同じ突出の仕方をしている。そして実際は両端が同じ制限酵素で切断されたDNA断片ができるので、図の中央に書いたような形の断片ができる。これも180°回転させても同じ形の断片となることが確認できる。両端が3´粘着末端の断片は英語の「S」、両端が5´粘着末端の断片は英語の「Z」に似ている。そして「S」「Z」とも180°回転させても同じ形になることからもイメージできるのではないでしょうか。
(180°回転対称は数学では「点対称」ともいいます。)
図の下は、同じ図を枠線なしで→だけで描いたものです。この描き方になれると更に簡便に思考できるように
なります。
【6塩基認識配列が異なる制限酵素の切断面でも、5´粘着末端どうし(3´粘着末端どうし)で、粘着末端が同じならば結合できる】
多くの制限酵素は6塩基認識です。そして粘着末端(突出部)はその2~5塩基目となります。すると、結合が可能な3´粘着末端どうし(5´粘着末端どうし)で、1・6番目塩基が異なっていると6塩基配列は異なる制限酵素が認識するが、2~5塩基目が共通の場合、次のようなことがおきます。本設問の制限酵素YとZはともに5´粘着末端であり、2~5番目が共通で、この例の1つです。
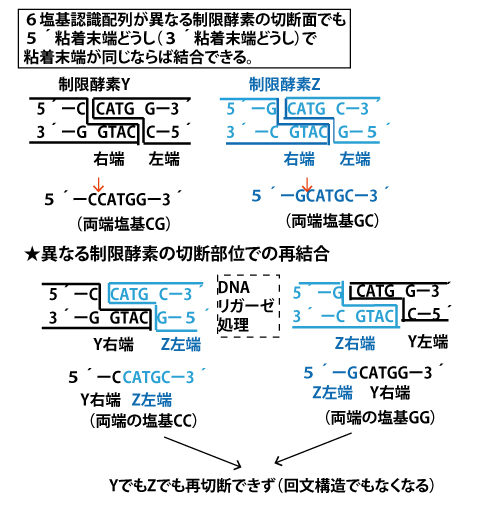
Y切断面とZ切断面の一本鎖突出部分4塩基(粘着末端)は同じなので「のりしろ」が同じようなもので、DNAリガーゼという「のり」に相当する酵素で処理すると、粘着末端どうしの塩基対が相補的に水素結合し、鎖が連結します。Y右端とZ左端、Z右端とY左端の2種類の結合が可能になります。(図中の一本鎖の塩基表記は、これまで述べてきたように、1本鎖だけでも考える練習のための表記です、)
更に面白いことに、この異なる制限酵素の切断面が結合したDNAの連結部は1・6塩基目に関し元の制限酵素2種が認識する配列と異なる(回文構造にもなっていない)新たな配列となっているため、Y・Zで再切断しようとしてもできません。
この現象は共通の粘着末端部分(2~5塩基)は除外し、1・6塩基目だけ取り出して考えるとわかりやすいです。
1・6塩基目に関し、Y認識は「CG」、Z認識は「GC」、異なる断片が結合した場合は「CC」あるいは「GG」となり、Y・Zでは再切断されないことがわかります。
さて最後に本設問に戻ると、Xのみが3´粘着末端、YとZは5´粘着末端で、YとZは粘着末端部分が共通(CATG)なのでYとZの断面どうしが結合が可能です。
よってプラスミド挿入部分の左(X切断面)に結合可能なのはXのみ、右(Z切断面)に結合可能かのはYとZ。
よって断片aとcが可能。よって![]()
(配点5点、正答率31%、部分正答率![]() (cのみ)(2点)27.7%、
(cのみ)(2点)27.7%、![]()
(aのみ)(1点)4.6%)
問2
細胞における情報伝達と遺伝子発現の流れを確認しておこう。図を上から順にみてほしい。
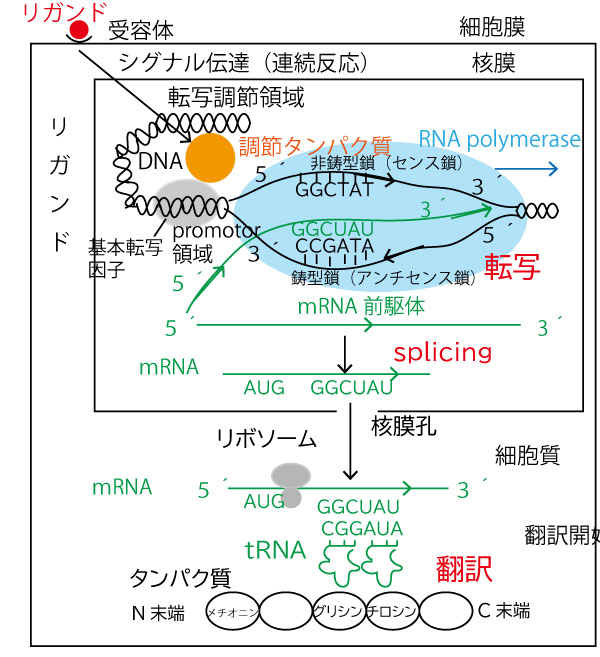
他の細胞からの体液(血液など)を通じて細胞表面に到達した刺激物質をリガンドと総称する。ホルモンなどがその一例である。それが細胞膜表面の
受容体に受け止められと、細胞室内(核外)にシグナル伝達という連続反応が起き、その情報を受け継いだ調節タンパク質が核内に移行し、遺伝子上流の転写調節領域(設問図のア)に結合する。すると核内にもともと存在していた基本転写因子との協調により、その近傍にDNAからmRNAの転写を始める「RNA polymerase」が結合する。基本転写因子やRNA polymeraseが結合するDNA部分をpromotor(設問図のイ、転写開始点直前であることからわかる)という。
RNA polymeraseはDNA二重らせんをほどき、片側(鋳型鎖)の3´→5´方向(図では青い→方向)に進みながら、それと相補的な塩基配列を持つmRNA前駆体を5´→3´方向に合成していく。この過程を転写という。この時、DNAの反対側の鎖(非鋳型鎖)はだたほどかれるだけであり、こちらの鎖ではmRNA前駆体合成は行っていない。
転写されたmRNA前駆体に不要な部分(intron)を除去し、必要な部分(exon)だけをつなぎ合わせるsplicingが行われ、短いmRNAとなる。
(なおこのDNA・mRNAには鎖に全て塩基配列があるが、図では簡略のため、9塩基のみを示した。)
mRNAは細胞質に移動し、リボソームがmRNAの5´側から少し進んだ翻訳開始点(本設問図のウ)(AUG)からその塩基配列情報を読み取り、3塩基ごとにそれに相補的な塩基を持つtRNAを引き寄せる。tRNAは特定のアミノ酸を運び、そのアミノ酸どうしがペプチド結合しタンパク質が合成される。これを翻訳という。なおmRNAの5´→3´方向の順に合成されるアミノ酸配列では
N末端(アミノ基末端)→C末端(カルボキシ基末端)となり、これがタンパク質合成の順番の向きになる。
よって答は![]()
(配点3点、正答率40.3%)
問3
GFP(green fluorescent protein、緑色蛍光タンパク質)とは下村脩(しもむらおさむ)氏が発見したオワンクラゲ由来の蛍光物質で、分子生物学で細胞内の動きを追跡したいタンパク質に遺伝子にGFPの遺伝子を融合した遺伝子を組み込むと、その物質の細胞内での位置や動きが蛍光で観察できる(細胞イメージング技術)。下村氏はこの業績で2008年ノーベル化学賞を受賞した。
細胞骨格(cytoskeleton)とは細胞内で構造の維持や物質や小器官の移動に関与する繊維状物質であり、チューブリンでできた微小管、中間径フィラメント、アクチンフィラメントの3種類がある。「微小管」という名前であるが、この3種の中で最も太く、アクチンフィラメントが最も細い。
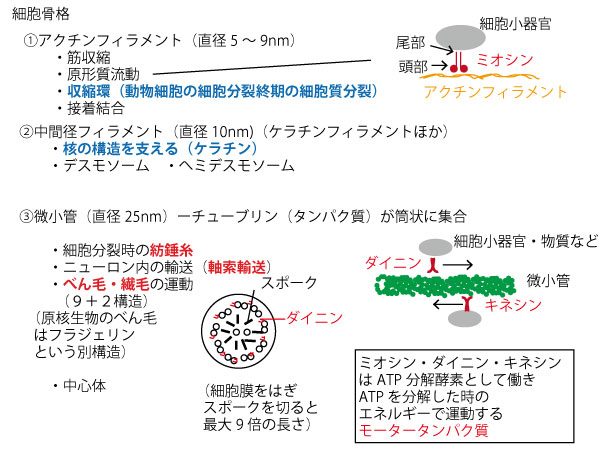
図にあるように動物細胞分裂の収縮環(図d)はアクチンフィラメント、核の構造(図e)を支えるのは中間径フィラメント(ケラチン)である。核には、蛍光が観察される微小管は、紡錘糸(f)・べん毛(g)・軸索(h)にあり、そこにGFPの蛍光も観察される。よって、![]()
(配点3点、正答率15.8%)

